4.6 The inner hair cells
The inner hair cells (IHCs) are the cochlea’s actual transducers. There are about 3,500 of them, arranged in a single row along the entire length of the basilar membrane, each one connected to about 10–30 fibers of the auditory nerve. Whatever leaves the cochlea — what the brain ever gets to hear — is encoded in the firing of the spiral ganglion neurons that the inner hair cells drive. The IHCs are the bottleneck.
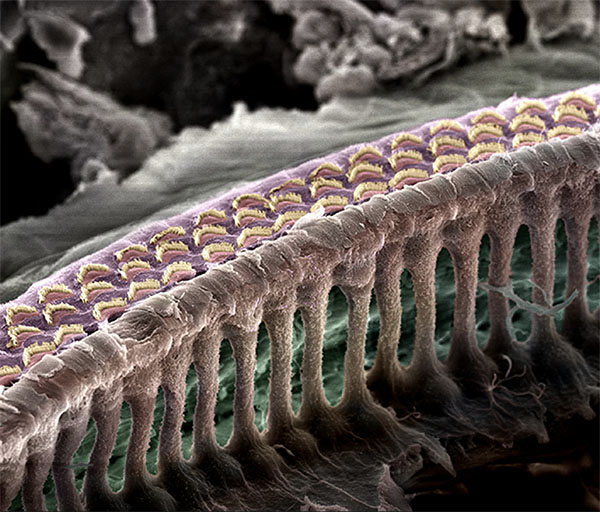
Stereocilia, tip links, and MET channels
Each IHC sits with its body in the organ of Corti and a tuft of about 100 stereocilia — actin-filled rod-like protrusions — emerging from its apical surface into the scala media (the endolymph-filled chamber). The stereocilia are arranged in a graded array, taller on one side, shorter on the other, with each row connected to the next by tiny molecular threads called tip links. When the basilar membrane moves, the resulting fluid motion shears the tectorial membrane against the stereocilia tuft, and the whole bundle pivots — bending toward the tallest row in one direction and toward the shortest row in the other.
This deflection is the mechanical event that the cell transduces. On each stereocilium near the top of the bundle sits a mechanically gated ion channel — the MET (mechanoelectrical transduction) channel. The tip link connects the top of one stereocilium to the side of its taller neighbor. When the bundle bends toward the tall side, the tip link stretches, which mechanically pulls the MET channel open. When it bends toward the short side, the link slackens and the channel closes. The channel is permeable to K⁺ (and Ca²⁺), and because the scala media is held at a +80 mV endocochlear potential relative to the cell interior at –60 mV, opening the channel drives a K⁺ current into the cell, depolarizing it.
The Boltzmann transduction curve
The relationship between bundle deflection and open probability is well-approximated by a Boltzmann function:
The width parameter is on the order of 25 nm. The operating point is biased so that at rest about 15–20% of the channels are open. This bias matters: it means small deflections in either direction produce open-probability changes (and hence voltage changes), and it means the input-output relation is asymmetric — positive deflections produce larger depolarizations than equally-large negative deflections produce hyperpolarizations.
▶ Derivation: the Boltzmann distribution from two-state thermodynamics Derivation
The MET channel can be in one of two states — open () or closed () — with a free-energy difference between them. From equilibrium statistical mechanics, the probability of finding the channel in state is
where is Boltzmann’s constant and is absolute temperature.
The free-energy difference depends on the mechanical force exerted on the channel via the tip link. For a channel with a “gating swing” — the distance the channel moves between closed and open states — pulling on the tip link with force contributes to (force times displacement, with the sign favoring the open state when ). And the tip-link force is, to first approximation, linear in bundle deflection: , where is the tip-link stiffness.
Substituting:
So
where is the deflection at which the channel is half-open and is the characteristic width of the transition. This is the Boltzmann sigmoid in the main text.
The numerical value nm corresponds, at body temperature, to a tip-link force of roughly per nm of deflection — which, combined with a few-nm gating swing, gives micro-piconewton-scale force changes per nm of deflection. The cell is reading mechanical forces at the limit of thermal fluctuation. ∎
The interactive below renders this transduction curve and animates it. Drag the deflection-amplitude slider to set how hard the bundle is being shaken (the visual rate is constant, decoupled from real frequency, so the motion stays watchable). The cursor on the curve marks the operating point; the lower trace shows the stimulus (gray, sinusoidal deflection) against the resulting receptor potential (black). At small amplitudes, the receptor potential is a small sinusoid centered on rest, mostly symmetric. At large amplitudes, the asymmetry of the Boltzmann produces rectification: the receptor potential develops a DC offset on top of its AC oscillation. The cell sits in a more depolarized state on average. This DC component is what allows the IHC to follow envelopes and to drive transmitter release in a graded way; the AC component is what carries the precise timing information that the auditory nerve will phase-lock to in 6.3.
- current deflection
- 60.0 nm
- open probability
- 0.900
- V_receptor
- -53.2 mV
The ribbon synapse
The depolarization opens voltage-gated Ca²⁺ channels in the IHC’s basal pole, near where the cell synapses onto auditory-nerve fibers. The resulting Ca²⁺ influx triggers vesicle release at ribbon synapses — a specialized synaptic structure unique to ribbon-bearing sensory cells (the IHC, the retinal photoreceptor, the IHCs of birds and fish). Ribbon synapses can release vesicles continuously, faithfully, at rates approaching 1000 per second per ribbon, which is what gives the auditory system its extraordinary timing precision. The released glutamate binds AMPA receptors on the postsynaptic auditory-nerve terminal and triggers spike generation. We will pick this up in detail in movement 6.
IHC vs OHC, the asymmetry
A note on the asymmetry of inner vs outer hair cells, since it has been hovering at the edge of the discussion since 4.1. They are anatomically similar but functionally orthogonal. The IHC is a transducer that says nothing about the mechanics of the membrane it sits on — it just reports what the membrane is doing. The OHC is a mechanical actuator that contributes to the motion of the membrane and ordinarily does not contribute much to the brain’s spike trains. The synaptic count is the most telling number: about 95% of the afferent fibers in the auditory nerve come from inner hair cells; only about 5% come from outer hair cells, and even those are not what carries the auditory signal — they may serve a feedback or modulatory role we still do not fully understand. The vast majority of the efferent fibers running from the brainstem back into the cochlea, on the other hand, terminate on outer hair cells, where they modulate the gain of the cochlear amplifier. The brain talks back to the OHCs. It mostly only listens to the IHCs.