The cochlea is, above all, an acoustic instrument. It does Fourier analysis with water and bone. The reason we are about to spend a section on its geometry is not that the geometry is intrinsically interesting — though it is — but that every detail of the geometry has an acoustic job. The fluids are wave media. The membranes are mechanical resonators. The hair cells are transducers reading out the displacement of one of those resonators. When “Hey Dr. Miles!” arrives at the oval window, it is going to fill this whole space; and what comes out the other end will look nothing like what went in.
So this section is anatomy, but anatomy with a purpose. As we lay out each piece — the three fluid-filled chambers, the membranes between them, the cells that read the result — try to keep one question in mind: what is this for, mechanically? In 4.2 we begin to answer that quantitatively; for now we just want the stage.
The cochlea is a spiral about 9 mm across; uncoiled, it would run roughly 35 mm. It is buried in the densest bone in the body, the petrous part of the temporal bone, which is part of why most casual descriptions of it leave the geometry hand-waved. To do the physics later, we need the shape clean.
The structure that matters is what the cochlea looks like in cross-section — what you would see if you sliced perpendicular to its coiled axis at any one place along its length. The cross-section is qualitatively the same everywhere, but the dimensions of its parts change as you walk from the base, near the stapes, to the apex, the far tip of the spiral. That 35 mm walk is the variable we will care about more than any other; from a mechanical standpoint it is the coordinate axis along which everything interesting happens.
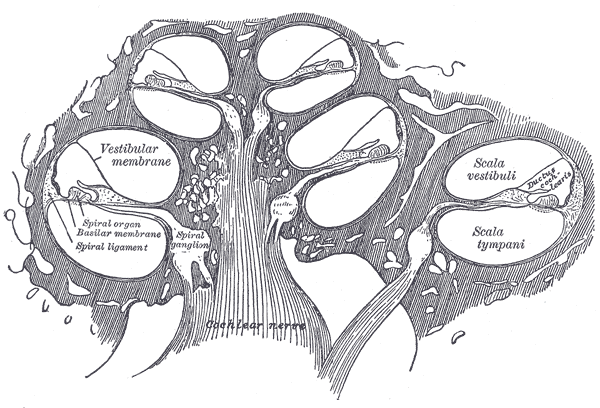
The image below is the original anatomical reference for the cross-section, from the 1918 edition of Gray’s Anatomy. It looks slightly bewildering on first inspection, which is honest — the structure is dense — but the next few paragraphs will give us names for every region you see.
Gray's plate 928: cross-section of one turn of the cochlea. Drawn by Henry Vandyke Carter.Henry Gray & H. V. Carter · public domain · Wikimedia Commons
In cross-section the cochlea is three fluid-filled tubes stacked vertically. These are fluid tubes, which matters: the perilymph and endolymph are essentially incompressible, and they conduct mechanical disturbances the way water does, not the way air does. The wave physics in the cochlea is the wave physics of pressure variations in nearly-incompressible liquid, driven by a small piston (the stapes) and constrained by membrane and bone. From top to bottom, the three chambers are:
The scala vestibuli, filled with perilymph — a fluid whose ionic composition is similar to extracellular fluid (high Na⁺, low K⁺). This is the chamber the stapes pushes into.
The scala media, filled with endolymph — an unusual fluid that, ionically, looks more like the inside of a cell (high K⁺, low Na⁺). The K⁺-rich endolymph is held at a positive resting potential of about +80 mV relative to perilymph; this endocochlear potential is part of what powers transduction in 4.6.
The scala tympani, again perilymph.
The membrane separating vestibuli from media is Reissner’s membrane, a thin sheet whose mechanical role is debated; for the wave physics we are about to do, it acts as essentially transparent and we will treat it that way. The membrane between media and tympani is the basilar membrane, and this is the protagonist of the next five sections. It is the resonator. The whole story of frequency analysis in the cochlea is the story of how the basilar membrane moves.
Sitting on top of the basilar membrane, inside scala media, is a structure called the organ of Corti. It is a row of cells — sensory cells, supporting cells, and an overhanging gelatinous strip called the tectorial membrane — arranged in a specific architecture that converts membrane motion into electrical signals. There are two flavors of sensory cell, and the difference between them is central. The inner hair cells, of which there are about 3,500 in a single column, are the actual transducers — they sense the motion of the basilar membrane and send it to the brain. The outer hair cells, of which there are about 12,000 in three columns, are not really transducers in the usual sense; they are active mechanical elements that sharpen the tuning of the basilar membrane beyond what passive mechanics allows. Section 4.5 develops this mechanism; its consequences run through movement 9.
The interactive below is the cross-section in schematic form. Hover any element, on the diagram or in the legend, to bring it forward. The rest of the movement assumes familiarity with the location of the basilar membrane and the organ of Corti.
Scala vestibuli(perilymph) — Upper chamber. Receives the stapes input at the base of the cochlea.
Reissner's membrane — Thin sheet separating vestibuli from media. Largely passive in the wave physics; we will treat it as transparent.
Scala media(endolymph) — Middle chamber. Contains the organ of Corti, bathed in an unusual K⁺-rich fluid that establishes the endocochlear potential.
Basilar membrane — The resonator. The protagonist of the next five sections.
Organ of Corti — Sits on the basilar membrane. Contains the sensory hair cells and their supporting cells.
Tectorial membrane — Overhangs the hair cells. Couples basilar-membrane motion to the stereocilia of the hair cells.
Inner hair cells — ~3,500 cells in a single column. The actual transducers; their spikes are what leave the cochlea.
Outer hair cells — ~12,000 cells in three columns. Active mechanical elements; they sharpen the tuning of the membrane in a way passive mechanics cannot.
Scala tympani(perilymph) — Lower chamber. Terminates at the round window, which bulges out to accommodate the incompressible fluid pushed in at the oval window.
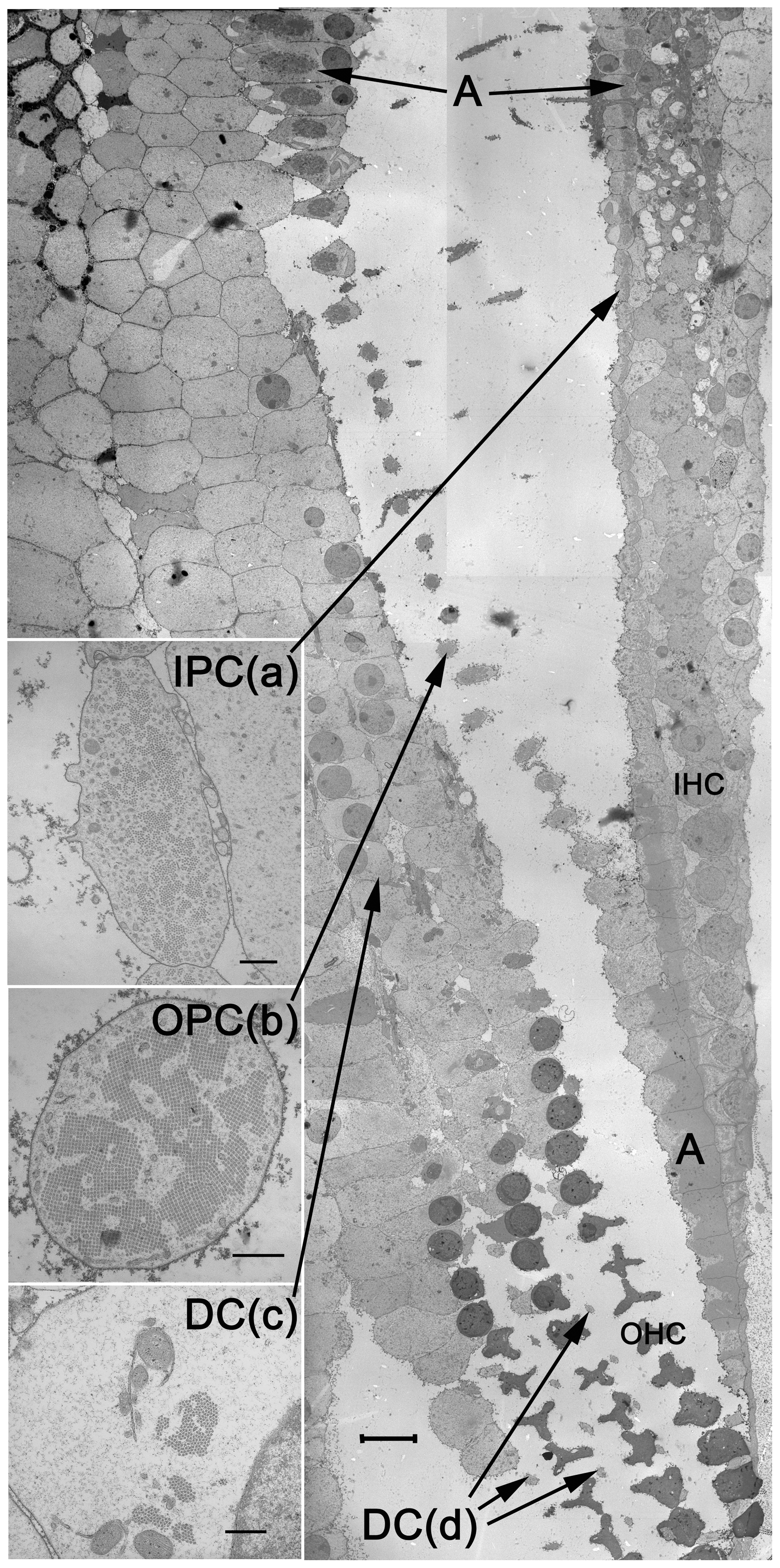
A schematic is an abstraction; the image below shows the cells themselves. The image below is a real scanning electron micrograph through the organ of Corti, sectioned at the level of the basilar membrane. You are looking at the pillar cells (IPC, OPC) that triangulate the tunnel of Corti, the Deiters’ cells (DC) that cradle the outer hair cells, and the inner and outer hair cells (IHC, OHC) themselves. The reticular lamina sits like a tiled roof across the top of all of this.
The organ of Corti at the level of the basilar membrane, in scanning electron microscopy. Inner pillar cells (IPC), outer pillar cells (OPC), Deiters' cells (DC), inner hair cells (IHC), and outer hair cells (OHC) are labelled.Zetes, Tolomeo & Holley, 2012 · CC BY 4.0 · Wikimedia Commons
Three more pieces of geometry are worth pinning down before we move on:
The oval window is the input port at the base of the scala vestibuli. The footplate of the stapes pushes fluid into the cochlea here. It is the piston that drives the whole system.
The round window is a flexible membrane at the base of the scala tympani. It exists for a banal but crucial reason: the inner-ear fluid is incompressible, so when the stapes pushes in at the oval window, the round window has to bulge out to give the fluid somewhere to go. Without it, the stapes would have nothing to push against and the cochlea would be inert.
At the apex of the spiral, scala vestibuli and scala tympani meet through a small opening called the helicotrema, which short-circuits any DC pressure imbalance between the two perilymphatic chambers. We will use this in 4.3 when we draw the boundary conditions for the traveling wave.
The geometry is now in place. What we don’t yet have is any quantitative claim about how the basilar membrane behaves. That is the next question, and where the math finally starts.
⏳The history— Alfonso Corti and the organ that bears his name
Alfonso Corti, an Italian anatomist working in Würzburg, published the first detailed description of the sensory epithelium of the cochlea in 1851. Working with hardened preparations and early compound microscopes, he identified the rows of pillar cells, the arch they form (the tunnel of Corti), and the hair-bearing cells on either side. He did not know what the cells did — the connection between hair cells and hearing would take another half-century — but the anatomy was precise enough that the structure still carries his name.
Corti left science shortly afterward and returned to manage his family’s estate; the work that made him famous was essentially his only major publication.